


光谱和光度特性概述

本文概述了荧光蛋白及其光谱特性。随着 20 世纪 50 年代末荧光蛋白的发现,荧光显微技术发生了巨大变化。它始于 O. Shimomura 和来自水母(Aequorea victoria)的绿色荧光蛋白(GFP)[1]。后来出现了数百种 GFP 突变体,发出的荧光从蓝色到红色不等。来自安氏动物(海葵和珊瑚)的荧光蛋白则将发射转移到了远红外[2]。蛋白质种类繁多,科学家们可以根据自己的需要选择zui合适的荧光标签。


什么是绿色荧光蛋白(GFP)?
GFP(绿色荧光蛋白)或其变体的光谱特性取决于构成发色团的氨基酸结构(图 1)[2]。这可能是位于 65-67 位的三个氨基酸或靠近该位置的残基(如 YFP)。除了有关发色团的主要突变外,还进行了其他定点诱变研究,以改善蛋白质成熟和在异源细胞系统中表达等其他因素(如密码子使用、蛋白质在生理温度下的折叠)。请注意,维多利亚甲虫是一种相对原始的外温海洋生物,没有调节体温的生理系统。

图 1:GFP 的分子结构。
尽管 GFP 因其亮度和高光稳定性而成为zuishouhuanying的荧光粉之一,但它也有两个主要缺点:对 pH 值有一定的敏感性和轻微的二聚倾向。二聚化或寡聚化是许多 FP 的一个问题。它们相互凝集的倾向会产生假象或误解融合蛋白的位置和功能。不过,科学家们也找到了一些解决这一问题的办法。在关键位置(F223R、L221K 和 A206K)的非极性氨基酸被亲水性氨基酸取代后,二聚化程度降低[3]。所有能改善光谱和实际特性的基因改变都被归纳为"增强型"FP。
就 wtGFP 而言,增强后的 EGFP(增强型 GFP)在 488 纳米波长处只有一个激发峰,而不是以前在 395 纳米波长和 475 纳米波长处的复杂吸收光谱[2,3]。Roger Heim、Douglas Prasher 和 Roger Tsien 开发的第一种 wtGFP 突变体(S65T 突变体)的亮度是原来的五倍,而且成熟时间更短[3]。加上另一个突变体(F64L)在 37°C 温度下的成熟效率更高,这对于观察活细胞的人来说具有重要作用。
蓝宝石(Sapphire)[3]是一个非常有趣的 GFP 变体,它的斯托克斯位移最大。靠近发色团的一个位置发生了突变(T203I),导致激发最大值变为 399 纳米,发射最大值变为 511 纳米。这就是 112 纳米的斯托克斯偏移。Emerald 是另一种 GFP 改造品,它能提高光稳定性和亮度,并能在哺乳动物细胞中更有效地折叠[3]。
所有绿色荧光蛋白都具有相对较高的亮度,而蓝色荧光蛋白在显微应用中通常会降低发射强度。尽管如此,由于具有其他光谱特性,它们仍被用于光学检测。EBFP(增强型蓝色荧光蛋白)是通过对 wtGFP 进行几轮突变而构建的[3]。第一个突变(Y66H)使发射峰从绿色变为蓝色。随后更多的突变产生了一种激发最大波长为 380 纳米、发射最大波长为 448 纳米的蛋白质。这些光谱特性使其成为 EGFP 在 FRET 显微镜下的搭档。最近,Azurite,SBFP2和 EBFP2 等蓝色荧光蛋白具有更高的量子产率和更好的光稳定性[3]。EBFP 的后继者是一种名为 Sirius 的蛋白质,这种蛋白质对 pH 值的耐受性非常高(在 pH 值 3 到 9 之间都很稳定),而且是迄今为止发射波长最短的荧光蛋白,因此很受欢迎[3]。
第二类 "蓝色" GFP 变体是由青色荧光蛋白 (CFPs)形成的[3]。用色氨酸取代酪氨酸(Y66W)并进一步改变基因,可产生亮度和光稳定性更好的荧光色素。这种 ECFP 在 433/445 纳米和 475/503 纳米具有双峰激发和发射光谱。亮度仅为 EGFP 的 40%。Cerulean 是 ECFP 的一个重要变体,它具有更高的消光系数和量子产率[3]。它的亮度是 ECFP 的 1.5 倍,被用作 YFP 的 FRET 伴侣。
GFP 的突变并不直接改变发色团三个中心氨基酸中的一个,因此出现了黄色荧光蛋白(YFP)[3]。YFP 在 203 位有一个共同的苏氨酸,该位置被一个酪氨酸(T203Y)取代。该氨基酸是 β 管的一部分,靠近发色团。与 GFP 相比,EYFP 的激发和发射特性已转向更长的波长,其激发和发射最大值分别为 514 纳米和 527 纳米(EYFP)。EYFP 的一个特点是对 pH 值敏感。在 pH 值为 6.5 时,EYFP 只有约 50%的荧光,但这并不总是一个缺点。在测量 pH 值(如囊泡、内体等)时,EYFP 可用作指示剂。有趣的是,进一步的突变(Q69M)产生了更好的酸稳定性并显著提高了亮度(比 EGFP 亮 75%)。与 EGFP 相比,这种蛋白质的光稳定性仍然较差,因此被称为Citrine[3]。另一种 YFP 突变体(F46L)的成熟速度大大加快,耐酸碱性也有所提高。这种蛋白质被命名为 Venus,是 Cerulean 的一种常见 FRET 受体[3]。
来自类动物的荧光蛋白质
如上图所示,来自维多利亚水母的大多数荧光蛋白发出的光谱为蓝色至黄色。红色荧光蛋白则不见踪影。俄罗斯科学家谢尔盖-卢基扬诺夫(Sergey Lukyanov)发现了拟水螅中的荧光蛋白,tianbu了这一空白[4]。与其他荧光蛋白相比,红色荧光蛋白(RFPs)具有很大的优势,因为在光谱的红色部分,细胞中的自发荧光要少得多[3]。此外,它们能被较长的波长激发,这对活细胞更有利,因为较短波长的光对标本的损害更大。与水母蛋白相比,珊瑚蛋白的另一个普遍优势是它们能在 37°C 温度下高效成熟。维多利亚A. GFP及其衍生物必须经过基因改造才能以正确的方式折叠,而动物蛋白的成熟则无需分子工程。这可能是由于其生物群落的水温较高。
在拟水生动物中发现的第一种 FPs,也是目前使用最多的 FPs 之一是DsRed[3]。其名称来源于海葵 Discosoma striata。DsRed 的激发最大波长为 558 nm,发射峰值为 583 nm。然而,当其结构信息公布后,最初的兴奋就停滞不前了。DsRed 的成熟速度比水母 FP 慢得多,并且有一个中间发色团阶段。这一阶段发出的光为绿色光谱,会与其他 FP 重叠。GFP 部分已经提到,DsRed 还有另一个问题。海葵红色荧光蛋白是一种强制性四聚体,容易形成寡聚体。这可能导致对融合蛋白的位置和功能产生误解。一般来说,安氏无脊椎动物的 FPs 与维多利亚无脊椎动物的 FPs 具有相似的结构。发色团隐藏在大小为 4 nm x 3 nm(高 x 直径)的 β 管状结构中。不同之处在于安氏囊虫的 β-桶状结构更像椭圆形(图 2)。
在 GFP "进化" 的同时,研究人员开始对原始 DsRed 进行改造,以克服其结构缺陷。第二代 DsRed--DsRed2--减少了寡聚体的形成趋势,加快了成熟速度,最大限度地减少了绿色发光的中间阶段[3]。进一步的诱变导致红色 FP wanquan失去了四聚体状态,但也丧失了部分量子产率(DsRed2 的 25%)。钱氏实验室的这项工作产生了第一个单体红色荧光蛋白,因此他们称之为 mRFP1[3]。
随后,以这种 mRFP1 为起点,产生了一组六种单体 FP,统称为 "mFruit"[3]。它们各自的名称源自其发射颜色:mHoneydew、mBanana、mOrange、mTangerine、mStrawberry 和 mCherry。mCherry 是这些 FPs 中最有用的一种,它在 610 纳米范围内发光,亮度是 EGFP 的 50%。
迄今为止最耀眼的 FP 是 mFruit 派系的追随者,名为 tdTomato[3]。在基因改变之前,dTomato 是一种强制性二聚体,但通过将两个二聚化伙伴放在一个分子中,避免了二聚化。两个 dTomato 单元通过 12 个氨基酸连接体耦合,形成串联二聚体 FP tdTomato,其发射最大值为 581 纳米,在光谱的最高区域具有光稳定性。
mPlum是所有 mFruit 蛋白中红色发射最深的一种,发射波长为 649 nm[3]。

图 2:DsRed 荧光蛋白的分子结构。
科学上使用的绿色荧光类动物蛋白数量非常少,这并不令人惊讶,因为我们已经有了zhongsuozhouzhi,使用方便的维多利亚虫 GFP。显然,没有人认为有必要建立一种新的绿色荧光蛋白。尽管如此,还是有一些新的绿色荧光蛋白出现了,比如石珊瑚中的一种明亮荧光蛋白--Azami Green[3]。有趣的是,它与 EGFP 的序列同源性不到 6%。
Katushka 是一种对深部组织成像具有重大影响的拟态荧光蛋白[3]。通过对来自 E. quadricolor 的 RFPs 进行诱变,发现 Katushka 是一种二聚体蛋白,在 635 纳米波长处具有最大发射亮度,是所有深红色荧光蛋白中亮度zuigao的。Katushka的单体形式被称为 mKate,后来被提供了更高亮度的 mKate2[3]。
总之,今天用于显微应用的所有荧光蛋白都来自原始海洋生物。表 1 列出了最重要的荧光蛋白及其相关光谱特性,如激发和发射最大值、光稳定性、量子产率和亮度。
展 望
一个非常有趣的故事是,人们发现了一种由脊椎动物表达的 FP。文昌鱼(Amphioxus)是一种小型鱼类海洋脊索动物,在其前端产生AmphiGFP[3]。对该 FP 的序列分析表明,它具有典型的 β-桶状结构,似乎与桡足类 Pontellina plumata(甲壳动物)的 CopGFP有关[5]。这一发现表明,荧光现象并不局限于原始无脊椎动物,在更高进化阶段的动物中也能发现。此外,这一发现还表明,荧光蛋白的发现、改造和增强过程仍在继续,这也是目前的热门研究课题。这一事实证明了荧光蛋白对当前和未来生命科学研究的重要性和巨大影响。
荧光蛋白质的光谱特性
Ex:峰值激发波长(纳米)
Em:峰值发射波长(纳米)
MW:分子量 QY:量子产率
BR:亮度;
消光系数 * 量子产率/1,000
PS:光稳定性;亮度达到 50%的时间(秒)
< 上下滑动查看全部 >
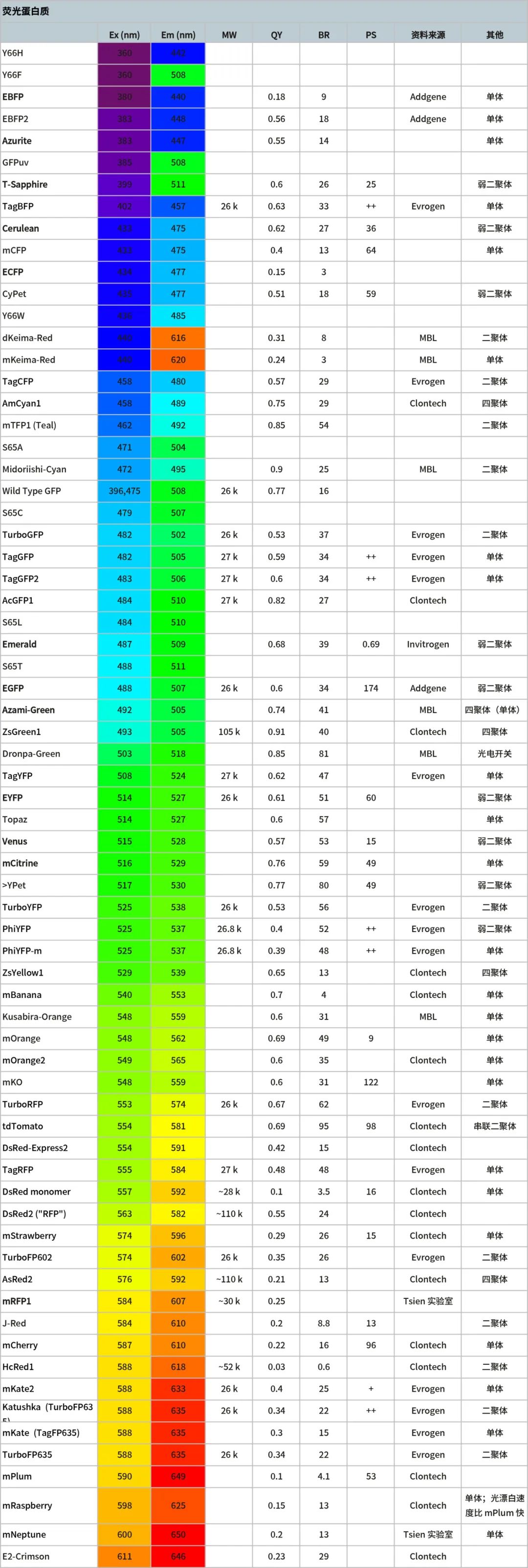
表 1:荧光蛋白的光谱特性 (数据来源为参与文献 6)。
相关产品

DMi 8gaoduan倒置显微镜平台

STELLARIS 5 & STELLARIS 8共聚焦显微镜平台

MICA全场景显微成像分析平台
参考文献:(上下滑动查看更多)
本文地址: https://www.xsyiq.com/44139.html
网站内容如侵犯了您的权益,请联系我们删除。
